
В одній хромосомі розміщено багато генів. Неалельні гени можуть міститися в одній, або в різних хромосомах. Якщо гени розташовані у різних хромосомах, вони будуть успадковуватися незалежно.
Американський генетик Томас Морган вивчав закономірності успадкування генів, розташованих в одній гомологічній хромосомі.

Томас Морган
В експериментах Морган використовував плодову мушку дрозофілу, що має важливі для генетиків якості: невибагливість, плодовитість, невелику кількість хромосом (чотири пари), безліч чітко виражених альтернативних ознак.


Дрозофіла та її хромосомний набір
У мухи дрозофіли гени, що відповідають за забарвлення тіла і довжину крил, розташовуються в одній хромосомі: домінантна ознака \(A\) зумовлює сіре тіло у дрозофіли, рецесивна \(a\) — чорне тіло; домінантна \(B\) — нормальні крила, \(b\) — короткі крила.
Морган проводив два типи аналізуючого схрещування. При схрещуванні рецесивної за обома ознаками самки (чорної з короткими крилами) з дигетерозиготним самцем (сірим з нормальними крилами) у потомстві з'явилося \(50\) % чорних мух з короткими крилами і \(50\) % мух з сірим тілом і нормальними крилами.
Отже, гени, розташовані в одній хромосомі, успадковувалися разом. Такі гени утворюють групу зчеплення.
Явище спільного успадкування генів, які містяться в одній хромосомі, Морган назвав зчепленим спадкуванням.
Інші результати було отримано, коли схрещували дигібридну самку (сіру з нормальними крилами) з гомозиготним рецесивним самцем (чорним з короткими крилами) . У цьому випадку потомство мало чотири фенотипи: \(41,5\) % — сірих мух з короткими крилами, \(41,5\) % — чорних з нормальними крилами, \(8,5\) % — сірих з нормальними крилами, \(8,5\) % — чорних з короткими крилами.
Морган прийшов до висновку, що зчеплення може бути неповним. Воно порушується в результаті кросинговеру — обміну ділянками між гомологічними хромосомами.
Кросинговер
У профазі \(I\) мейозу відбувається кон'югація гомологічних хромосом. Хромосоми зближуються, а потім починають розходитися і утворюють перехрест (хіазми). У процесі кон'югації між деякими дочірніми хроматидами є можливим обмін ділянками (кросинговер).
Кросинговер — обмін ділянками між гомологічними хромосомами, який відбувається у профазі \(I\) мейозу.
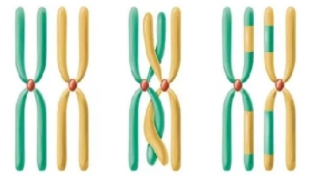
Кожна з утворених хроматид потрапляє в окрему гамету. У результаті кросинговеру утворюються кросоверні гамети, хромосоми яких містять нові комбінації генів.
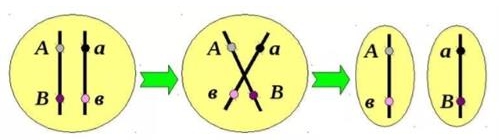
Організми, які виникають в результаті злиття кросоверних гамет, називають рекомбінантними або кросоверними.
Оскільки кросинговер відбувається не після кожної кон'югації, то і число кросоверних гамет є завжди меншим числа некросоверних гамет.
Морган довів, що частота кросинговеру між гомологічними хромосомами залежить від відстані між генами у хромосомі. Чим ця відстань є більшою, тим частіше відбувається кросинговер і тим частіше з'являються рекомбінантні організми. Частота рекомбінацій (кросинговеру) визначається у відсотках до загального числа нащадків
Частота рекомбінацій (кросинговеру) = число рекомбінантів : загальне число нащадків × \(100\)%.
Ця величина показує відносну відстань між зчепленими генами у хромосомі. За одиницю відстані між генами приймають \(1\) морганід (М) (\(1\) % кросинговеру), або відсоток появи рекомбінантних особин.
Частота рекомбінацій (кросинговеру) = число рекомбінантів : загальне число нащадків × \(100\)%.
Ця величина показує відносну відстань між зчепленими генами у хромосомі. За одиницю відстані між генами приймають \(1\) морганід (М) (\(1\) % кросинговеру), або відсоток появи рекомбінантних особин.
У дослідженнях Т. Моргана кількість кросоверних організмів дрозофіл становить \(8,5\) + \(8,5\) = \(17\) %, а некросоверних — \(83\) %. Отже відстань між генами дорівнює Aa і Bb \(17\) сМ.